1Division of Biotechnology, IIHR-ICAR, Hessaraghatta Lake Post, Hessaraghatta, Bangalore, India-560089
2Division of Plant Pathology, IIHR-ICAR, Hessaraghatta Lake Post, Hessaraghatta, Bangalore, India-560089
3Department of Biotechnology, Centre for post-graduate studies, Jain university, Bangalore, India
Corresponding author Email: prasu8050655414@gmail.com
Article Publishing History
Received: 29/12/2018
Accepted After Revision: 26/03/2019
RNA interference is a sequence dependent gene silencing mechanism with huge potential in agriculture. Virus disease
management has become a daunting task due to the lack of availability of resistant cultivars. In this scenario, RNAi mediated viral resistance has acquired importance. RNAi can be employed to silence the viral genes conferring virus resistance. Transformation of the host with an RNAi vector synthesizing hairpin RNA is a prerequisite. It is highly desirable to develop multiple virus resistance due to the fact that, viruses counter the host RNAi by employing silencing suppressors there by leading to resistance breakdown. This can be addressed by targeting all the commonly infecting viruses along with viral slicing suppressors. This increases the complexity of RNAi vector construction as multiple fragments are involved to develop an ihp construct. OE(Overlap-Extension)-PCR is the method of choice due to its several advantages such as no extraneous sequences are incorporated unlike as in restriction based fragment joining and it is also simple and economical compared to other methods. In this experiment, we demonstrate the use of OE-PCR in the construction of an intron containing hairpin RNA synthesizing vector targeting seven genes from four viruses viz. that commonly to tomato and chilli for imparting multiple virus resistance viz. capsicum chlorosis virus, groundnut bud necrosis virus, cucumber mosaic virus and chilli veinal mottle virus. Seven viral gene fragments were joined together in to 1.64kb fragment by OE-PCR. The complete stretch was assembled into the pBI121 and mobilized in to Agrobacterium for transformation.
Overlap Pcr, Rna Interference, Virus Resistance, Agrobacterium
Babu K. P, Manamohan M, Reddy M. K. Overlap Extension PCR to Anneal Multiple DNA Fragments for High-Throughput Double Stranded Rnai Vector Construction. Biosc.Biotech.Res.Comm. 2019;12(1).
Babu K. P, Manamohan M, Reddy M. K. Overlap Extension PCR to Anneal Multiple DNA Fragments for High-Throughput Double Stranded Rnai Vector Construction. Biosc.Biotech.Res.Comm. 2019;12(1). Available from: https://bit.ly/2EAkwT4
Copyright © Babu et al., This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-BY) https://creativecommns.org/licenses/by/4.0/, which permits unrestricted use distribution and reproduction in any medium, provide the original author and source are credited.
Introduction
Tomato (Solanum lycopersicum L.) and chilli (Capsicum annum) are commercially important vegetable crops grown globally. Virus management in these crops has been challenging in recent years due to intensive cultivation and spurt in the insect vectors population resulting in virus disease outbreak (Sastrosiswojo 1995; Umamaheswaran et al., 2003). Viral diseases caused by some major viruses like Tospovirus, Cucumovirus and Potyvirus results in immense economic loss in these crops productivity. Tospoviruses (capsicum chlorosis virus and groundnut bud necrosis virus) are negative single-stranded RNA virus, belongs to Bunyaviridae family and containing three genomic segments L, M and S, transmitted by thrips (Haan et al., 1990, 1991, Krishna Reddy et al., 2008). Cucumovirus (cucumber mosaic virus-CMV) is a positive sense single-stranded RNA virus belonging to family Bromoviridae (Palukaitis et al., 1992). Potyvirus (ChiVMV-chilli veinal mottle virus), belongs to the family Potyviridae, is a single molecule which is linear, positive-sense, single-stranded RNA (ssRNA). Potyvirus and cucumovirus are transmitted by insect vector aphids in a non-persistent manner (Berger et al., 2005). Current virus disease management involves prophylactics, roguing and insect vector control. Non availability of the resistance gene pools in most of the cultivated crops poses a serious hurdle in resistant breeding. In this regard RNA interference is highly promising in imparting virus resistance (Simon-Mateo and García 2011, Duan et al., 2012; Li et al., 2106; Jia et al., 2017).
RNA interference is a natural antiviral defense system, degrades the RNA molecules in a sequence-specific manner and it is activated in response to double-stranded RNA formed during viral replication (Voinnet et al., 2001). RNAi mediated gene silencing involves, formation of double-stranded RNA, so called “hairpin” RNA (hpRNA), formed by annealing of the reverse complementary sequence at one half of the RNA, which is recognized by and activates plant RNAi machinery leading to the degradation the viral RNA, finally plants recovering from viral infection. The expression of virus-specifi c dsRNA as hairpin structures renders the plants resistant against virus infection (Bonfi m et al., 2007). In plants, intron-mediated hairpin RNA (ihpRNA) by incorporating an intron as a spacer sequence between the sense and antisense fragments results in higher gene silencing efficiency in the transgenic plants (Wesley et al., 2001). Moreover, fi eld level virus resistance is achieved by targeting multiple viruses in a single chimeric construct (Bucher et al., 2006; Zhang et al., 2011; Thu et al., 2016; Hameed et al., 2017).
Therefore, a cost effi cient, high-throughput system for designing and making of ihpRNA constructs is in great demand. To achieve broad spectrum resistance, there is a need to target multiple virus genes, to join these fragments the selection of restriction enzymes may not be possible in case multiple fragment splicing due to the limited choice and incompatibility of restriction enzymes and also the introduction of extraneous nontarget sequences might interfere with the silencing efficiency. Due to these diffi culties, OE-PCR is the method of choice to join multiple fragments.
OE-PCR is a technique used to synthesize artificialgenes, joining one or more DNA segments together to form a new genetic combination (Li et al., 2008) The OE-PCR was fi rst described by (Higuchi et al., 1988). The basic principle of OE-PCR is that individual DNA fragments are designed to have a short stretch of their 3’ ends to be complimentary and these overlaps are extended in PCR to splice the two fragments in to one (Harton et al., 1989), the initial PCR fragments then generate overlap gene fragments that can be used as new templates for another PCR to form a full length fragment (Heckman et al., 2007; Wäneskog and Bjerling 2014; Hussain and Chong 2016).
Targeting multiple viruses is important in aiming for RNAi based robust and durable resistance. In this study, RNAi gene construct against broad spectrum of viruses was designed based on the conserved sequences from four different viruses commonly infesting tomato and chilli viz., Capsicum chlorosis virus-nucleocapsid (CaCV-N), Groundnut bud necrosis virus-Nucleocapsid and Non-structural proteins (GBNV-N and NSs), Cucumber mosaic virus-2b and Coat protein (CMV-2b and CP) and Chilli veinal mottle virus-helper component proteinase and coat protein (ChiVMV HC-Pro and CP). We used OE-PCR to join all seven fragments in a single stretch to construct hpRNAi-MVR binary vector for plant transformation to impart multiple virus resistance in tomato and chilli. This is the fi rst report by using OE-PCR for joining seven gene fragments in ihpRNA construct for plant virus resistance.
Materials and Methods
Selection of target gene sequences for dsRNA construct
Based on the literature survey, we have selected major virus diseases affecting tomato and chilli in Southern India, viz. GBNV, CaCV, CMV, ChiVMV diseases have caused a drastic yield loss in India. To develop broad spectrum ihpRNA vector, we have selected the above four viruses, from these 7 crucial genes were selected to express hairpin RNA in transgenic plants. The sequences of these genes were retrieved from GenBank at the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/BLAST/). BioEdit-multiple sequence alignment using Clustal W (Hall 1999) was carried out and highly conserved regions were selected for dsRNA design from the alignment of about 100- 150 viral sequences reported. The selected conserved regions of CaCV-N gene (255bp), GBNV-N gene (255bp) and GBNV-NSs gene (261bp) from Tospovirus, CMV 2b (232bp) and CMV CP gene (214bp) from Cucumovirus and ChiVMV HC-Pro (209bp) and ChiVMV-CP (221bp) from Potyvirus respectively. The detailed GenBank accession for the selected genes included in the (table 1). Off-target effect minimization was carried out using dscheck and Genome BLAST analysis to select highly conserved regions.
Overlap Primer designing and amplifi cation of desired gene fragments The clones of viral genes were obtained from the Division of Plant pathology, ICAR-IIHR, Bengaluru. The overlap primers were designed for the individual fragments of fragment S1, S2, S3, S4, S5 and S6 and it generates overhang region of 10-15bp to join the next fragment. No restriction enzymes were incorporated between these fragments. Restriction sites of BamH1/Sac1 and SexA1 were added in 5’ region of S1 fragment and 3’ region of S7 fragment to form ihpRNA construct forming sense: Intron: Antisense orientation. All individual fragments were amplifi ed using PCR. All PCR reactions were carried out in 25μl reaction contains 50ng of template, the PCR mix contains 1x Phusion HF buffer 3% DMSO, 200 μM DNTPs, 0.5 μM PCR primers and 1U Phusion® High- Fidelity DNA Polymerase (NEB, USA). The PCR conditions were as follows: initial denaturation for 30 seconds at 98 °C, followed by 35 cycles of denaturation 10 seconds at 98 °C, 30 seconds at 55 °C annealing, 10 seconds at 72 °C extension and the fi nal extension 30 minutes at 72 °C.
The amplifi ed products were purifi ed using NucleoSpin ® Gel and PCR Clean-up kit (Macherey-Nagel, Germany), and ligated into PTZ57R/T cloning vector by using T4 DNA ligase (Thermo scientifi c, USA). The ligated products were transformed in to E.coli (DH5ᵅ) using heat shock method. The transformed clones were sequenced by Sanger Sequencing method (Eurofi ns Genomics India Pvt Ltd, Bangalore). The confi rmed clones were used further for overlap PCR.
Joining of fragments by Overlap PCR
Three rounds of overlap PCR were carried out to join all 7 fragments. The PCR parameters for the modifi ed Overlap PCR is as follows: PCR was carried out in a 50μl reaction contains 50ng of purifi ed individual fragments, the PCR mix contains 1x Phusion HF buffer 3% DMSO, 200 μM DNTPs, 0.5 μM PCR primers and 1U Phusion® High-Fidelity DNA Polymerase (NEB, USA) the PCR mix contains no primers. The overlap PCR as follows: initial denaturation for 1 minute at 98 °C followed by extension at 72 °C for 15 minutes, pause/hold the programme at 72 °C or place the tubes in ice, then add the end primers and resume the programme followed by 35 cycles of denaturation for 10 seconds at 98 °C, annealing for 30 seconds at 60 °C, extension for 30s/kb at 72 °C and fi nal extension for 30 minutes at 72 °C. The above PCR conditions were followed to join all fragments.
In First round of overlap PCR, the fragments of S1+S2 were joined by using F1 and R2 primers, S4+S5 fragments were joined by using F4 and R5 primers and the fragments of S6+S7 were joined by using F6 and R7 primers respectively. An aliquot of all post PCR products were confi rmed on agarose gel, the confi rmed products were purifi ed by NucleoSpin® Gel and PCR Clean-up kit (Macherey-Nagel, Germany). The Second round overlap PCR was carried out by joining the fragments of S1+S2 with S3 using F1 and R3 primers and fragments of S4+S5 with S6+S7 were joined using F4 and R7 primers. An aliquot of post PCR products was used for confi rmation of amplifi cation and the rest was used for purifi cation. Likewise, the fi nal round PCR was continued by joining the fragments of S1+S2+S3 with S4+S5+S6+S7 using F1 and R7 primers (Fig 1 and 2). The BamH1/Sac1 restriction sites were incorporated in the F1 primer and SexA1 restriction site were incorporated in R7 primer, used to form Sense and Antisense fragments. A complete stretch 1.647kb fragment was purifi ed and ligated in to PTZ57R/T cloning vector (Thermo scientifi c, USA) using T4 DNA ligase (Thermo scientifi c, USA) and the ligated product was transformed in E.coli (DH5 cells). Plasmids were isolated using alkaline lysis method and three clones were sequenced for confi rmation of the fragments joined by OE-PCR.
The complete stretch containing all three fragments of 7 genes in sense and antisense orientation interspersed by PDS intron was released using BamH1 and SacI restriction sites, this fragment was purifi ed and ligated into the binary vector pBI121 between CaMV 35S promoter and NOS terminator using BamH1 and SacI sites. The ligation of sense-intron-antisense fragments in pBI121 binary vector were confi rmed by restriction digestion using BamHI and SacI. The resulting plasmid was designated as hpRNAi-MVR. The RNAi expression cassette (hpRNAi-MVR) was electroporated into Agrobacterium tumefaciens strain EHA105, the electroporated cells were grown on selectable antibiotics kanamycin 100mg l-1 and rifampicin 25mg l-1 containing LB media and the clones were confi rmed by PCR and Restriction digestion. Glycerol stocks were prepared for
the confirmed clones and stored in -80 °C for further use.

Results and Discussion
PCR amplifi cation of seven individual fragments by using proof reading Phusion DNA Polymerase yielded the expected amplicon sizes as assessed by agarose gel electrophoresis and sequencing. The expected amplicon sizes were CaCV-N gene 255bp, GBNV-NSs gene 261bp, GBNV-N gene 255bp, CMV-2b gene 232bp, CMV-CP gene 214bp, ChiVMV-HC pro gene 209bp and ChiVMVCP gene 221bp respectively.
Overlap extension PCR was carried in 3 successive rounds of PCR reactions to effi ciently complete the joining of all the seven fragments (Fig. 1).
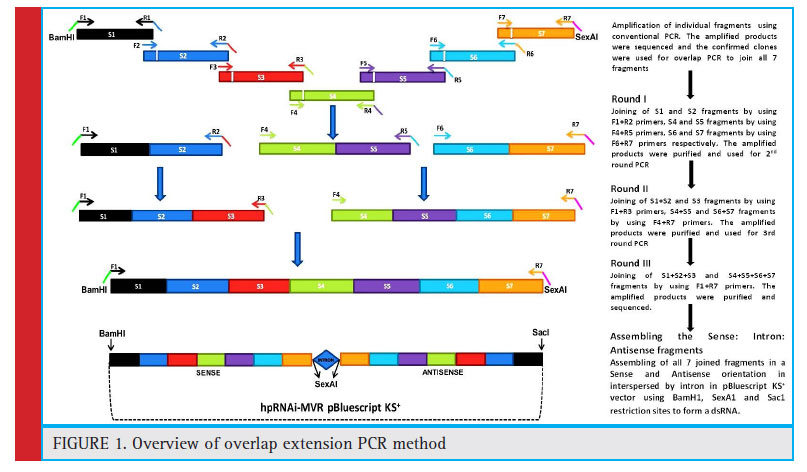 |
Figure 1: Overview of overlap extension PCR method |
In first round of OE-PCR, the S1+S2, S4+S5 and S6+S7 fragments were successfully joined separately with the expected size if about 516bp, 446bp and 430bp. The amplifi ed PCR products were purified and these eluted products were subsequently used for next round of OE-PCR. In second round OE-PCR, S1+S2+S3 and S4+S5+S6+S7 were joined successfully and the fragment sizes were 771bp and 876bp. Third round OE-PCR, assembled all seven fragments by combining fragments obtained from the second round OE-PCR. The size of the final fragment was about 1647bp. Sequencing revealed that there were no errors or mutations/ mismatches after at splicing of all the 7 fragments (Fig. 2).
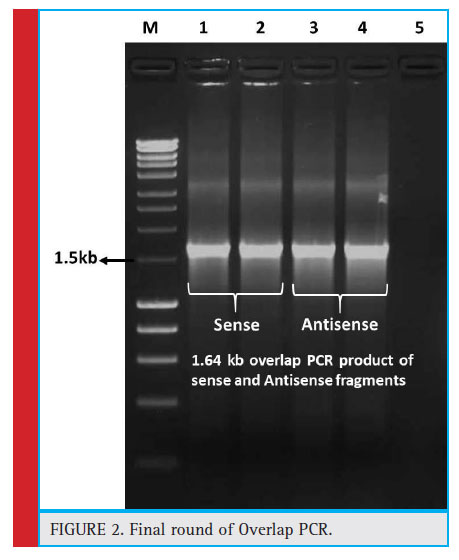 |
Figure 2: Final round of Overlap PCR. |
The amplicon represents the results of fi nal round overlap PCR, for sense fragment, BamH1 forward primer and SexA1 reverse primer (F1+R7) were used and for Antisense fragment Sac1 forward primer and SexA1 reverse primer (F1+R7) were used. The gel picture depicts that the expected amplicons of both sense and antisense fragments. M. HyperLadder 1kb; 1, 2. Final overlap PCR product of Sense fragment of about 1.64kb; 3,5. Final overlap PCR product of Antisense fragment of about 1.64kb; 5. No template control (NTC).
To form dsRNA, the complete stretch of 1.647kb fragment in sense and antisense orientation with the intervening 0.25kb of pdsintron were ligated and cloned, the plasmids were purifi ed and the orientation of sense, intron and antisense fragments was confirmed by respective restriction digestion and release of fragment of expected size (Fig. 3).
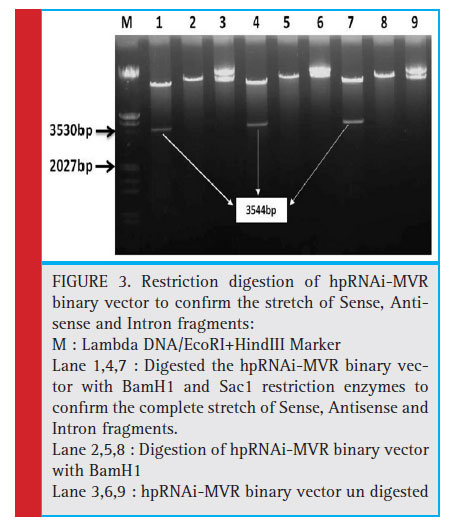
Discussion
Biotic stresses like viral diseases pose serious threat to crop production and estimated economic loss ranks second only to the other pathogens (Simon-Mateo and Garcia 2011). Effi cient virus diseases control measures are not available and current virus management strategy in most cases are limited to insect vector control and roguing. Development of virus disease resistant lines is highly desirable and RNAi has a direct bearing in the development of genetically engineered virus resistant crops (Chicas and Macino 2001). Imparting virus resistance has been proven effective in various plant species by using hpRNAi constructs (Ammara et al., 2015; Jia et al., 2017; Kumar et al., 2017).
Towards this, in this experiment we have developed a gene construct targeting multiple viruses commonly infecting two important Solancaeous crops, chilli and tomato by employing OE-PCR. Targeting single virus may not be sufficient as most often, the viral infections are mixed under field conditions, to overcome this problem, and also to minimize the resistance breakdown by a mechanism employed by viruses to overcome host induced viral gene silencing, known as “suppression of silencing” targeting multiple gene segments of virus including the respective silencing suppressors could therefore result in more durable and robust virus resistance to broad spectrum of virus infections under field conditions. In this regard, we have selected viral silencing suppressors like NSs gene for Tospovirus (Goswami et al., 2012; Zhai et al., 201 4), 2b gene for Cucumovirus(Du et al., 2014; Nemes et al., 2014) a nd HC-Pro gene for Potyvirus (Torres-Barcelo et al., 2010; Sahana et al., 2014) for broad spectrum virus resistance. The viral suppressors plays an important role, it protects the viral genome against RNA silencing and acts as RNA silencing suppressor (Lakatos et al., 2006). Hc-Pro gene from potyvirus blocks PTGS at tissue level, CMV 2b from cucumovirus prevents systemic silencing (Brigneti et al., 1998). NSs gene from tospovirus regulates the suppression of the viral movement and host defense mechanism (Lokesh et al., 2010). Many researchers have reported resistance to multiple virus infections (Bucher et al., 2006; Zhang et al., 2011; Thu et al., 2016; Hameed et al., 2017). In this experiment, we have selected seven fragments to target seven viral genes, and this renders the development of gene construct a tedious and complex task as the commonly employed restriction digestion and ligation of fragments gets unwieldy as the number of fragments to be joined is more. Thus, we have employed OEPCR successfully to accomplish the assembly of hpRNAi construct.
Splicing of two fragments of DNA, referred as overlap extension PCR established by (Yolov et al., 1990), this method has many advantages like joining of many fragments of about 10.8kb using pfu proofreading polymerase (Shevchuk et al., 2004). Majority of commercial kits and long PCR protocols are mainly based on Taq polymerase thus cannot be used for overlap extension PCR, because Taq leaves a single A-overhang at 3’ region. This leads to disrupt the primer binding at overlap regions during overlap extension PCR (Sambrook et al., 2001). The overlap extension PCR has many applications like, joining of 26 synthetic oligos of 54nt of about 969bp with 5 rounds of sequential overlap extension PCR (Zhang et al., 2013). Many modifi ed methods of OE-PCR have been applied in many genetic engineering areas, such as fusion of long length DNA fragments (Fujii et al., 2013; Engel et al., 2013), assembly of longer gene fragments (Benders et al., 2010; You et al., 2012).
Here, we developed a broad spectrum, multiple virus resistant hpRNAi-MVR vector by targeting 7 genes from 4 common virus groups for both tomato and chilli crops viz., CaCV N gene, PBNV N and NSs genes from Tospovirus, CMV 2b and CP gene from cucumovirus and HCPro and Polyprotein from ChiVMV. We have successfully employed the overlap extension PCR method to splice multiple gene fragments without using any restriction sites and adaptors etc. as the addition of non-specific sequences in the gene construct would have affected the RNAi effi cacy due to its sequence dependence. The use of OE-PCR is highly desirable in the development of RNAi constructs involving splicing of multiple fragments. Simplifi ed and efficient gene construct assembly for RNAi would aid in the development of virus resistant crops in avoiding crop yield loss.
Conclusion
Seamless joining of multiple fragments to create single fragment without using any restriction sites or ligase is a critical step in RNAi. Here, we demonstrated the application of overlap extension PCR to join seven viral gene fragments to develop hpRNAi-MVR construct useful for imparting virus disease resistance in both tomato and chilli. Virus diseases drastically reduce the crop yield and quality and are diffi cult to manage and only the long term solution to this is to develop virus resistant crop lines. Development of broad spectrum hpRNAi construct for commonly infecting viruses is one of the important aspects. Here, we developed an hpRNAi-MVR vector that is useful in developing multiple virus resistant tomato and chilli employing OE-PCR. Demonstrate that OE-PCR is a method of choice in RNAi gene construct for virus resistance as it is simple, fast and economical, can join multiple fragments in less time. This approach can be employed in developing gene constructs for durable, multiple virus resistance in other important crops.
Acknowledgment
We are thankful to ICAR-IIHR, which provided the facility to carry out this research work. This is the part of Ph.D. dissertation work of fi rst author.
References
Ammara U., Mansoor S., Saeed M., Amin I., Briddon R.W. and Al-Sadi A.M. (2015). RNA interference-based resistance in transgenic tomato plants against Tomato yellow leaf curl virus-Oman (TYLCV-OM) and its associated betasatellite. Virology journal, 12(1): 38.
Benders G.A., Noskov V.N., Denisova E.A., Lartigue C., Gibson D.G., Assad-Garcia N., Chuang R.Y., Carrera W., Moodie M., Algire M.A., Phan Q., Alperovich N., Vashee S., Merryman C., Venter J.C., Smith H.O., Glass J.I., Hutchison C.A. (2010). Cloning whole bacterial genomes in yeast. Nucleic Acids Research, 38 (8): 2558–2569.
Berger P.H., Adams M.J., Barnett O.W., Brunt A.A., Hammond J., Hill J.H., Jordan R.L., Kashiwazaki S., Rybicki E., Spence N., Stenger D.C., Ohki S.T., Uyeda I., van Zaayen A., Valkonen J., Vetten H.J Family potyviridae. In: Fauquet C.M., Mayo M.A., Maniloff J., Dessel Berger U., Ball L. (eds) (2005). Virus taxonomy eighth report of the international committee on taxonomy of virus. Elsevier, Academic Press, London, 385–396.
Bonfi m K., Faria J.C., Nogueira E.O., Mendes E.A. and Aragao F.J. (2007). RNAi-mediated resistance to Bean golden mosaic virus in genetically engineered common bean (Phaseolus vulgaris). Molecular Plant-Microbe Interactions, 20 (6): 717-726.
Brigneti G., Voinnet O., Li W.X., Ji L.H., Ding S.H. and Baulcombe D.C. (1998). Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO Journal, 17: 6739–6746.
Bucher E., Lohuis D., van Poppel P.M., Geerts-Dimitriadou C., Goldbach R. and Prins M. (2006). Multiple virus resistance at a high frequency using a single transgene construct. Journal of General Virology, 87 (12): 3697-3701.
Chicas A. and Macino G. (2001). Characteristics of post transcriptional gene silencing. EMBO reports, 2 (11): 992-996. De Haan P., Kormelink R., de Oliveira Resende R., Van Poelwijk F., Peters D. and Goldbach R. (1991). Tomato spotted wilt virus L RNA encodes a putative RNA polymerase. Journal of General Virology, 72 (9): 2207-2216.
De Haan P., Wagemakers L., Peters D. and Goldbach R. (1990). The S RNA segment of tomato spotted wilt virus has an ambisense character. Journal of General Virology, 71 (5): 1001- 1007.
Du Z., Chen A., Chen W., Liao Q., Zhang H., Bao Y., Roossinck M.J. and Carr J.P. (2014). Nuclear-cytoplasmic partitioning of the Cucumber mosaic virus 2b protein determines the balance between its roles as a virulence determinant and RNA silencing suppressor. Journal of virology, JVI-00284.
Duan C.G., Wang C.H. and Guo H.S. (2012). Application of RNA silencing to plant disease resistance. Silence, 3 (1): 5.
Engel H., Gutiérrez-Fernández J., Flückiger C., Martínez-Ripoll M., Mühlemann K., Hermoso J.A., Hilty M., Hathaway L.J. (2013) Heteroresistance to fosfomycin is predominant in Streptococcus pneumoniae and depends on the murA1 gene. Antimicrobial Agents, 57 (6): 2801–2808.
Fujii W., Kano K., Sugiura K., Naito K. (2013). Repeatable construction method for engineered zinc fi nger nuclease based on overlap extension PCR and TA-cloning. PloS one, 8 (3): e59801.
Goswami S., Sahana N., Pandey V., Doblas P., Jain R.K., Palukaitis P., Canto T. and Praveen S. (2012). Interference in plant defense and development by non-structural protein NSs of Groundnut bud necrosis virus. Virus research, 163 (1): 368- 373.
Hall T.A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic acids symposium series, 41 (41): 95-98.
Hameed A., Tahir M.N., Asad S., Bilal R., Van Eck J., Jander G. and Mansoor S. (2017). RNAi-mediated simultaneous resistance against three RNA viruses in potato. Molecular biotechnology, 59 (2-3): 73-83.
Heckman K.L., Pease L.R. (2007). Gene splicing and mutagenesis by PCR-driven overlap extension. Nat Protocols, 2: 924– 932.
Higuchi R. (1988). A general method of in vitro preparation and specifi c mutagenesis of DNA fragments: study of protein and DNA interactions. Nucleic Acids Research, 16: 7351–7367.
Horton R.M., Hunt H.D., Ho S.N., Pullen J.K. and Pease L.R. (1989). Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene, 77 (1): 61-68.
Hussain H., and Chong N.F.M. (2016). Combined overlap extension PCR method for improved site directed mutagenesis. BioMed research international (doi.org/10.1155/2016/8041532).
Jia R., Zhao H., Huang J., Kong H., Zhang Y., Guo J., Huang Q., Guo Y., Wei Q., Zuo J. and Zhu Y.J. (2017). Use of RNAi technology to develop a PRSV-resistant transgenic papaya. Scientifi c Reports, 7 (1): 12636.
Krishnareddy M., Rani R.U., Kumar K.A., Reddy K.M. and Pappu H.R. (2008). Capsicum chlorosis virus (Genus Tospovirus) infecting Chili pepper (Capsicum annuum) in India. Plant disease, 92 (10): 1469-1469.
Kumar S., Tanti B., Patil B.L., Mukherjee S.K., Sahoo L. (2017). RNAi-derived transgenic resistance to Mungbean yellow mosaic India virus in cowpea. PLoS ONE, 12 (10): e0186786.
Lakatos L., Csorba T., Pantaleo V., Chapman E.J., Carrington J.C., Liu Y.P., Dolja V.V., Calvino L.F., Lopez., Moya J.J. and Burgyan J. (2006). Small RNA binding is a common strategy to suppress RNA silencing by several viral suppressors. The EMBO journal, 25 (12): 2768-2780.
Li X., Qiu Y., Shen Y., Ding C., Liu P., Zhou J. and Ma Z. (2008).
Splicing together different regions of a gene by modifi ed polymerase chain reaction-based site-directed mutagenesis. Analytical biochemistry, 373 (2): 398-400.
Li L.I., Cheng G.U., Biao W.A., Tong Z.H., Yang L.E., DAI Y.H., Wen H.E., LIANG C., WANG X.F. (2016). RNAi-mediated transgenic rice resistance to Rice stripe virus. Journal of integrative agriculture, 15(11):2539-49.
Lokesh B., Rashmi P.R., Amruta B.S., Srisathiyanarayanan D., Murthy M.R. and Savithri H.S. (2010). NSs encoded by groundnut
bud necrosis virus is a bifunctional enzyme. PLoS One, 5 (3): e9757.
Manamohan M., Chandra G.S., Asokan R., Deepa H., Prakash M.N. and Kumar N.K. (2013). One-step DNA fragment assembly
for expressing intron-containing hairpin RNA in plants for gene silencing. Analytical biochemistry, 433 (2): 189-191.
Nemes K., Gellert A., Balazs E. and Salanki K. (2014). Alanine scanning of Cucumber mosaic virus (CMV) 2B protein identifies different positions for cell-to-cell movement and gene silencing suppressor activity. PloS one, 9 (11): e112095.
Palukaitis P., Roossinck M.J., Dietzgen R.G. and Francki R.I. (1992). Cucumber mosaic virus. In Advances in virus research. Academic Press, 41: 281-348.
Sahana N., Kaur H., Jain R.K., Palukaitis P., Canto T. and Praveen S. (2014). The asparagine residue in the FRNK box of potyviral helper-component protease is critical for its small RNA binding and subcellular localization. Journal of General Virology, 95 (5): 1167-1177.
Sambrook J. and Russel D.W. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Simon-Mateo C. and Garcia J.A. (2011). Antiviral strategies in plants based on RNA silencing. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms, 1809 (11): 722-731.
Thu L.T.M., Thuy V.T.X., Duc L.H., Son L.V., Ha C.H. and Mau C.H. (2016). RNAi-mediated resistance to SMV and BYMV in transgenic tobacco. Crop Breeding and Applied Biotechnology, 16 (3): 213-218.
Torres-Barcelo C., Daros J.A. and Elena S.F. (2010). HC-Pro hypo-and hyper suppressor mutants: differences in viral siRNA accumulation in vivo and siRNA binding activity in vitro. Archives of virology, 155 (2): 251-254.
Voinnet O. (2001). RNA silencing as a plant immune system against viruses. Trends Genetics, 17: 449–459.
Shevchuk N.A., Bryksin, A.V., Nusinovich, Y.A., Cabello, F.C., Sutherland, M., Ladisch S. (2004). Construction of long DNA molecules using long PCR-based fusion of several fragments simultaneously. Nucleic Acids Research, 32 (12): e19.
Waneskog M and Bjerling P. (2014). Multi-fragment sitedirected mutagenic overlap extension polymerase chain reaction as a competitive alternative to the enzymatic assembly method. Analytical biochemistry, 444: 32-37.
Wesley S.V., Helliwell, C.A., Smith, N.A., Wang, M.B., Rouse, D.T, et al. (2001). Construct design for effi cient, effective and highthroughput
gene silencing in plants. Plant Journal, 27: 581–590.
Yolov AA., Shabarova, ZA. (1990).Constructing DNA by polymerase recombination. Nucleic Acids Res, 18(13): 3983–3986.
You C, Zhang XZ, Zhang YHP. (2012). Simple cloning via direct transformation of PCR product (DNA Multimer) to Escherichia coli and Bacillus subtilis. Applied and environmental microbiology, 78(5): 1593–1595.
Zhai, Y., Bag, S., Mitter, N., Turina, M. and Pappu, H.R. (2014). Mutational analysis of two highly conserved motifs in the silencing suppressor encoded by tomato spotted wilt virus (genus Tospovirus, family Bunyaviridae). Archives of virology, 159(6): 1499-1504.
Zhang, X., Sato, S., Ye, X., Dorrance, A.E., Morris, T.J., Clemente, T.E. and Qu, F. (2011). Robust RNAi-based resistance to mixed infection of three viruses in soybean plants expressing separate short hairpins from a single transgene. Phytopathology, 101(11): 1264-1269.
Sastrosiswojo, S. (1995). Integrated pest management in vegetable production. Symposium on Small Scale vegetable production and Horticultural Economics in Developing Countries. ISHS Acta Horticulturae, 369.
Umamaheswaran, K., Jain, R. K., Bhat, A. I. and Ashwai, Y. S.9. (2003). Biological and molecular characterization of Tospovirus isolate from tomato and its relationship with other Tospovirus. Indian Phytopathology, 56: 168-173.

