1Parasitology and Microbiology Research Laboratory, Department of Zoology, University of Burdwan, India.
1Fishery and Ecotoxicology Research Laboratory, Department of Zoology, University of Burdwan, India.
Corresponding author email: microbiology@gmail.com
Article Publishing History
Received: 04/04/2021
Accepted After Revision: 28/06/2021
The coastal regions of Digha, West Bengal are endemic for lymphatic filariasis, especially Culex quinquefasciatus which is an established filarial vector there. Microorganisms in mosquito breeding sites is the key factor of either attraction or repellence of filarial vectors. The bacterial strains isolated from mosquito breeding sites were studied in this investigation. Culex quinquefasciatus plays a major role in transmitting lymphatic filariasis in the village areas of Digha, West Bengal. Water samples were collected from selected breeding habitats of Culex quinquefasciatus at the coastal villages of Digha. Five isolates (DX1, DX2, DX3, DX4 and DX5) were screened from the breeding habitats of Cx. quinquifasciatus, characterized and examined for oviposition bioassay. Test cup with the pure culture of Pseudomanas sp. DX5 was comparatively more attracted by gravid female Culex mosquitoes than other isolates in relation to oviposition.
Oviposition Activity Index (OAI) was 0.8. Phenotypic, biochemical and molecular characterization of Pseudomonas sp. DX5 was done. Pseudomanas sp. DX5 showed negativity to Gram stain. Organisms looked like rods without having any spores. The colonies were spherical, opaque, yellowish in colour. The strain was positive for catalase, oxidase, urease, H2S production and negative for indole production, methyl red and voges-proskauer test, citrate utilization and nitrate reduction test. It was sensitive to some standard antibiotics like levofloxacin, ofloxacin, ciprofloxacin, streptomycin and doxycycline but resistant against kanamycin, amoxycillin, nalidixic acid, ampicillin, chloramphenicol, gentamicin, tetracycline, vancomycin and erythromycin. Thus, elimination or control of the oviposition attractant bacterial strain of Pseudomanas sp. from breeding habitat water of filarial vector Cx. quinquifasciatus is an alternative strategy for filariasis management which is also a suitable topic of research in future.
Characterization, Culex Quinquefasciatus, Oviposition Activity Index, Pseudomonas Sp.14
Chatterjee S, Mandal A, Bag S, Sarkar B, Bhaduri R, Saha N. C . Isolation and Characterization of Pseudomonas sp. strain and its role as Oviposition Attractant of the filarial vector Culex quinquefasciatus. Biosc.Biotech.Res.Comm. 2021;14(2).
Chatterjee S, Mandal A, Bag S, Sarkar B, Bhaduri R, Saha N.C . Isolation and Characterization of Pseudomonas sp. strain and its role as Oviposition Attractant of the filarial vector Culex quinquefasciatus. Biosc.Biotech.Res.Comm. 2021;14(2). Available from: <a href=”https://bit.ly/2R4zQk9“>https://bit.ly/2R4zQk9</a>
Copyright © Chatterjee et al., This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-BY) https://creativecommns.org/licenses/by/4.0/, which permits unrestricted use distribution and reproduction in any medium, provide the original author and source are credited.
INTRODUCTION
Bancroftian filariasis is an important vector-borne disease transmitted by female Culex quinquefasciatus (Diptera: Culicidae) mosquitoes. The coastal belt of Digha in West Bengal, India is endemic for filariasis and Culex quinquefasciatus is the predominant mosquito vector at the coastal localities of Digha. The man-vector contact of Cx.quinquefasciatus is a key factor for high incidences of filarial infections in Digha (Azmi et al. 2015). The management of Culicidae is now difficult for the emergence of resistant vector mosquitoes to synthetic pesticides. Knowledge about the larval environments of Cx. quinquefasciatus is essential to control the mosquito vectors (Forattini, 1996).
The selection of suitable oviposition sites has a major role on progeny fitness, larval diversity and population dynamics and overall maternal reproductive fitness and success (Spencer et al. 2002). The mosquito oviposition and subsequently their site selection are influenced by multiple physical, chemical and environmental factors. These are also attractants, repellents or stimulating factors (Bentley and Day, 1989; Azmi et al. 2015; Mondal et al. 2019) .
It has been suggested by various workers that mosquito vectors have been influenced by bacterial semiochemicals (Leroy et al. 2011; Travanty et al. 2019). The oviposition attractants of mosquitoes include colour, odour, presence of semio-chemicals made by bacterial digestion or decomposition by bacteria of organic materials. The mosquito oviposition and subsequently their site selection are influenced by multiple physical, chemical and environmental factors. These are also attractants, repellents or stimulating factors (Bentley and Day, 1989; Mondal et al. 2019). It has been suggested by various workers that mosquito vectors have been influenced by bacterial semiochemicals (Leroy et al. 2011; Travanty et al. 2019).
Mosquitoes can interact with microbes in the breeding habitat water bodies in a variety of ways: as being the food items for the larvae, as being the symbionts, or as makers of kairomones (Merritt et al. 1992; Moreira et al. 2009; Shelomi 2019). Volatile compounds have been known to be associated with some specific bacterial strains that can enhance mosquito oviposition in the breeding sites of filarial vectors (Why and Walton 2020). Scanty literature is available in relation to the present theme of this study. Present study has been designed to isolate and characterize the oviposition attractant bacterial strains from the mosquito breeding habitats at the coastal regions of Digha, West Bengal.
MATERIAL AND METHODS
Samples were collected from varoius natural mosquito breeding habitat waterbodies in Champabani village of Digha from March 2019 to February 2020. Isolation, enumeration and pure culture of mosquito breeding habitat water bacteria were done on Nutrient Agar (Holt et al, 1994). The isolate was plated and cultured on MacConkey Agar (Himedia, India) and Pseudomonas Isolation Agar (Himedia, India). Morphological characters of bacterial colony (shape, size, colour, margin and opacity) were recorded (Technic et al. 1957).
Gram-staining of the isolates was done and cell morphology were checked under a Phase-Contrast Microscope. The surface topology of the bacteria was studied under Scanning Electron Microscope at various magnifications following standard protocols (Lacey 1997). Smears of the pure culture of the isolates were done on cover glasses and heat fixed for 1-2 sec. The smears were then fixed for 45 min in 2.5% glutaraldehyde solution.
The slides were then dehydrated by passing through 50%, 70% and 90% ethanol and finally with absolute alcohol for 10 min each. The samples were gold coated, scanned and photographed under Scanning Electron Microscope (ZEISS). In order to study the biochemical properties of the isolate, catalase test, indole test, methyl red test, voges-proskauer test, nitrate reduction test, citrate utilization test, urease test, oxidase test was performed following standard methodologies (Smibert and Krieg 1995; McClung 1985).
Sensitivity of the bacterial isolates to recommended doses of some commercially available standard antibiotics like kanamycin (30µg/disc), amoxycillin (10µg/disc), nalidixic acid (30µg/disc), ampicillin (10µg/disc), chloramphenicol (30 µg/disc), levofloxacin (5 µg/disc), gentamicin (50 µg/disc), ofloxacin (5 µg/disc), tetracycline (30 µg/disc), ciprofloxacin (5 µg/disc), vancomycin (30 µg/disc), rifampicin (5 µg/disc), erythromycin (15 µg/disc), streptomycin (10 µg/disc), and doxycycline (30 µg/disc) was recorded through disc diffusion technique following technique following standard methodology (Brown and Izundu 2004).
Pure culture of bacterial isolate (DX5) was inoculated in sterilized Nutrient Broth medium and incubated in a B.O.D. shaker incubator for 24h at 37 ± 2°C. Liquid bacterial culture of 1.8 mL of was centrifuged for 30 sec at 10000g to get the pellets and the genomic DNA was extracted by using DNeasy Ultra Clean Microbial Kit (Qiagen) following manufacturer`s instructions. Then amplification of nearly ~1.5 kb rDNA fragment of bacterial genomic DNA was extracted by using 27F forward and 1492R primer by polymerase chain reaction (PCR) technique (Brown and Izundu 2004).
Sequences of purified PCR products were obtained through universal bacterial primers in a DNA sequences. External service was taken for this purpose. Sequenced data were aligned in ClustalW software. As per the research by Jukes et al. (1969), evolutionary distances were calculated. Phylogenetic tree was built following standard method (Tamura et al. 2007). Larvae of Culex quinquefasciatus were collected from the breeding habitats in the coastal areas of Digha and the mosquito colonies were maintained at the Microbiology and Parasitology Research Laboratory, The University of Burdwan.
Mosquito larvae were reared in a water filled plastic bowls having sufficient yeast powder and dog biscuits in an acceptable ratio (1:1) in the laboratory conditions (29± 3 °C, 75-85% RH). Adult mosquitoes were released in wooden cages (30cm × 30cm × 30cm). Cotton soaked in 10 percent glucose solution was supplied to adult mosquitoes and adult female mosquitoes were regularly blood-fed to Wistar albino rat for egg development (Tamura et al. 2007).
The pure cultured bacterial isolates were discretely added to 100 ml Nutrient Broth and incubated at 30±1ºC for 72 hrs in a B.O.D shaker incubator to obtain a density of 105 cfu /ml. Sterilized Nutrient Broth media without-having any bacterial inoculum was taken as control. Ten adult gravid female Culex quinquefasciatus mosquitoes were released in each rearing cage (30cm x 30cm x 30cm). They were given two options for laying eggs. First cup was filled with 95 ml of sterile distilled water to which 5 ml (105 cfu/ml) of bacterial suspension was added.
The second cup filled with 100 ml of sterile distilled water without having any bacterial suspension served as the control. Separate set of experiment were conducted for each and every bacterial isolate. Ten replications were made for each treatment, using a fresh female Cx. quinquefasciatus for each treatment. Oviposition Activity Index (OAI) was determined by the formula: OAI = (Nt – Nc) / (Nt + Nc) [Nt= number of eggs laid in test cups; Nc = number of eggs in control cups] (Kramer and Mulla 1979). The entire experimental set-up was maintained in an environmental chamber at a temperature of 28±2ºC and relative humidity of 80±5% under 12:12h (light: dark) photoperiod (Kramer and Mulla 1979; Tamura et al. 2007).
Table 1. Number of eggs laid in test cups and control cups and Ovipositional Activity Index (OAI) in response to the suspensions of bacterial isolates.
| Types of water provided for oviposition in dual choice bioassay | No. of eggs laid in test cup (Mean ± S.E) | No. of eggs laid in control cup (Mean ± S.E) | Oviposition Activity Index (OAI) |
| (95mL of sterile distilled water + 5 mL of DX1) vs. 100mL of sterile distilled water
|
103±1.76 | 37±2.02 | 0.47 |
| (95mL of sterile distilled water + 5 mL of DX2) vs. 100mL of sterile distilled water
|
77 ± 3.21 | 37 ± 3.28 | 0.35 |
| (95mL of sterile distilled water + 5 mL of DX3) vs. 100mL of sterile distilled water
|
125.0 ±1.52 | 68±2.08 | 0.29 |
| (95mL of sterile distilled water + 5 mL of DX4) vs. 100mL of sterile distilled water
|
150.0±2.33 | 50±1.7 | 0.5 |
| (95mL of sterile distilled water + 5 mL of DX5) vs. 100mL of sterile distilled water | 383 ± 3.21 | 42.33 ± 2.33 | 0.8 |
| (95mL of sterile distilled water + 5 mL of sterile Nutrient Broth vs. 100mL of sterile distilled water)
|
63±2.64 | 38±1.7 | 0.24 |
RESULTS AND DISCUSSION
Five morphologically distinct bacterial colonies (DX1, DX2, DX3, DX4 and DX5) were screened from the breeding habitats of filarial vector Culex quinquefasciatus. Number of egg rafts in treated and control test cups has been shown in Table 1.
Gravid mosquitoes showed comparatively higher oviposition attractancy to the test cup having Pseudomanas sp. DX5 than other isolates. Oviposition Activity Index (OAI) was 0.8. The bacterial colonies of the isolate DX5 were spherical, opaque, yellowish in colour in Nutrient Agar. Bluish green coloured colony was developed in Pseudomonas Isolation Agar. The bacterial isolate DX5 was rod shaped without having any spore (Table 2).
Table 2. Cultural, phenotypic and biochemical properties of the bacterial isolate DX5
| Character | Observation |
| Colony character | Spherical, 2.5 mm, opaque, elevated, non- consistent, yellowish green |
| Vegetative cell | Rod shaped |
| Spore | Non spore forming |
| Gram stain | Negative |
| Biochemical tests | |
| Catalase | + |
| Indole test | – |
| Methyl red test | – |
| Voges–Proskauer test | – |
| Nitrate reduction test | – |
| Urease production test | + |
| Citrate test | – |
| Oxidase test | + |
| H2S production test | + |
| Antibiotic sensitivity Sensitive (lg/disc) | Doxycycline, streptomycin, levofloxacin, ofloxacin, ciprofloxacin, rifampicin |
| Resistant | Ampicillin, tetracycline, kanamycin, gentamycin, vancomycin, nalidixic acid, chloramphenicol |
Plate 1: Scanning Electron Micrograph of Bacterial isolate DX5
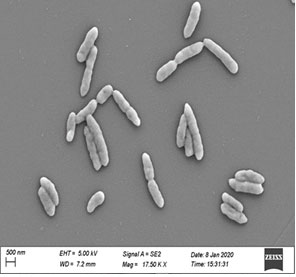
The Scanning Electron Micrograph of the bacterial isolate DX5 has been shown in Plate 1. The bacterial isolate (DX5) showed positive results for catalase, oxidase, urease, H2S production test and negative results for indole production, methyl red and voges-proskauer test, nitrate reduction test and citrate utilization (Table 2). The bacterial strain was sensitive to the recommended doses of levofloxacin, ofloxacin, ciprofloxacin, streptomycin and doxycycline but resistant against kanamycin, amoxycillin, nalidixic acid, ampicillin, chloramphenicol, gentamicin, tetracycline, vancomycin and erythromycin (Table 2).
The16S rRNA partial gene sequence of the bacterial isolate DX5 has been submitted in the NCBI GenBank with an accession number assigned as MW513479. Nucleotide base composition (mol%) has been depicted in (Fig 1).
Figure 1: Nucleotide composition of 16S ribosomal RNA gene sequence of bacterial isolate DX5
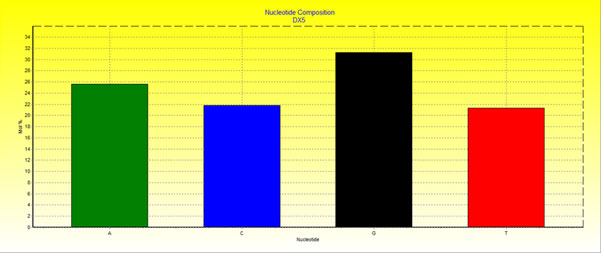
The values of AT content and GC content of the bacterial isolate DX5 were 46.92% and 53.08% respectively (Fig 1). Fingerprint of 16S rRNA partial gene sequence of Pseudomanas sp. DX5 has been depicted in (Fig 2).
Figure 2: Fingerprint of 16S rRNA gene sequences of bacterial isolate DX5
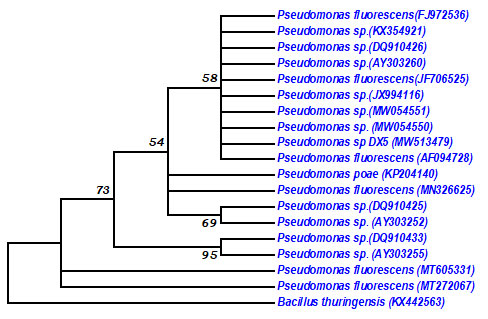
The Phylogenetic tree revealed that Pseudomonas sp. DX5 branched with Pseudomonas fluorescens (FJ972536) and Pseudomonas fluorescens (JF706525) having 58% bootstrap value (Fig 3).
Figure 3: Neighbour joining tree based on 16S rRNA gene sequence of Pseudomonas sp.DX5 with similar 16S rRNA gene sequences retrieved from NCBI and RDP database.
Coastal regions of Digha, West Bengal, India has been considered as filaria endemic region (Chandra et al. 2007; Azmi et al. 2015). Culex quinquefasciatus, the established filarial vector was the leading house-frequenting mosquito species in the coastal regions of Digha, West Bengal. The anthropophilic nature of Cx quinquefasciatusis was dependable factor for increasing intensity of filarial transmission in coastal areas of Digha (Azmi et al. 2015).
Recent observations reported by various workers have revealed that some bacterial strains could act as larval food, midgut flora, and its metabolites as important oviposition attractants and/or stimulants in mosquitoes. There are two key reasons for oviposition behaviour of a female mosquitoes. Gravid female mosquitoes must be induced for oviposition-by-oviposition media having specific bacterial suspension and also by some chemical inducers (Dethier et al. 1960). Oviposition attractancy depends on the composition and concentration of the microbial organisms found in breeding habitat water (Godwin et al. 2021).
Some strains of Acetobacter, Pseudomonas, Klebsiella, Gluconobacter, and Enterobacter spp. isolated from the insect orders Isoptera, Homoptera, Heteroptera, Coleoptera, Hymenoptera, and Diptera had important roles in larval development (Trexler et al. 2003; Roy et al. 2010). Microorganisms are responsible for the decomposition of detritus and release of volatile secondary metabolites. Gravid female Cx. quinquefasciatus mosquitoes were attracted to such habitats and were stimulated to lay eggs and Pseudomonas spp.
are considered as resident bacterial flora of different mosquito breeding habitats (Kennedy 1942; Gerhardt 1959; Ikeshoji 1968; Hosokawa et al. 2006, Rajagopal 2009). The oviposition bioassay clearly indicated that bacterial isolate DX5 (Pseudomonas sp.) served as an oviposition attractant for the filarial vector Culex quequinfasciatus. Now, the control of this oviposition attractant bacterial strains by some eco-friendly biocontrol agents like plant extracts and plant derived oils would certainly illuminate a new and alternative strategy of vector control.
CONCLUSION
The current study clearly showed that the strain of Pseudomonas sp. had a high oviposition attractancy index in relation to the oviposition of filarial vectors in the breeding habitats occurring at the coastal areas of Digha. Biocontrol or environmental management of the oviposition attractant bacterial strains in the mosquito breeding habitats of the coastal areas of Digha, West Bengal would be considered as an alternative strategy of vector control.
ACKNOWLEDGEMENTS
The authors are grateful to Department of Science and Technology and Biotechnology, Government of West Bengal for financial assistance for this work.
Conflict of Interests There was no conflict among the interests of the participating authors.
REFERENCES
Azmi, S. A., Das, S. and Chatterjee, S.N. (2015). Seasonal prevalence and blood meal analysis of filarial vector Culex quinquefasciatus in coastal areas of Digha, West Bengal, India. Journal of vector borne diseases, 52: 252.
Bentley, M. D. and Day, J. F. (1989). Chemical Ecology and Behavioral Aspects of Mosquito Oviposition. Annual Review of Entomology, 34: 401-421.
Brown, P. D. and Izundu, A. (2004). Antibiotic resistance in clinical isolates of Pseudomonas aeruginosa in Jamaica. Revista Panamericana de Salud Pública, 16: 125-130.
Chandra, G., Chatterjee, S. N., Das, S. and Sarkar, N. (2007). Lymphatic filariasis in the coastal areas of Digha, West Bengal, India. Tropical Doctor, 37: 136-139.
Dethier, V. G., Browne, B. L. and Smith, C. N. (1960). The Designation of Chemicals in Terms of the Responses They Elicit from Insects1. Journal of Economic Entomology, 53: 134-136.
Forattini, O. P. (1996). Culicidologia médica: identificação, biologia, epidemiologia Vol. 2, Edusp.
Gerhardt, R. (1959). The influence of soil fermentation on oviposition site selection by mosquitoes. Mosquito News, 19.
Godwin, U. O., Ifeoma, U. H., Barnabas, E.N., Chidozie, A. H., Winie, D. U. and Ogechi, K. Q. (2021). Bacteria and fungi profile of mosquito oviposition sites in Eziobodo, Owerri West LGA, Imo State.
Holt, J.G. Krieg, N.R., Sneath, P.H.A., Stanley, J.T., and Williams, S.T. (1994). Bergey’s Manual of Determinative Bacteriology (9th ed.), Williams & Wilkins, Co., Baltimore.
Hosokawa, T., Kikuchi, Y., Nikoh, N., Shimada, M. and Fukatsu, T. (2006). Strict host-symbiont cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biology, 4: e337.
Ikeshoji, T. (1968). Studies on Mosquito Attractants and Stimulants : V. Gas-Chromatographic Separation of the Attractants for Oviposition of Culex pipiens fatigans from the Field Water. Applied Entomology and Zoology, 3: 176-188.
Jukes, T. H., Cantor, C. R. and Munro, H. (1969). Mammalian protein metabolism.
Kennedy, J. S. (1942). On water-finding and oviposition by captive mosquitoes. Bulletin of Entomological Research, 32: 279-301.
Kramer, W. L. and Mulla, M. S. (1979). Oviposition attractants and repellents of mosquitoes: oviposition responses of Culex mosquitoes to organic infusions. Environmental Entomology, 8: 1111-1117.
Lacey, L. A. (1997). Preface. Manual of Techniques in Insect Pathology. Elsevier.
Leroy, P. D., Sabri, A., Verheggen, F. J., Francis, F., Thonart, P. and Haubruge, E. (2011). The semiochemically mediated interactions between bacteria and insects. Chemoecology, 21: 113-122.
Mcclung, L. (1985). Bergey’s Manual of Systematic Bacteriology, Volume 1.: Edited by Noel R. Krieg. The Williams & Wilkins Co., Baltimore, 1984, 964 pp. $80.00. International Journal of Systematic and Evolutionary Microbiology, 35: 408-408.
Merritt, R. W., Dadd, R. and Walker, E. D. (1992). Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annual review of entomology, 37: 349-374.
Mondal, R., Devi, N. P. and Jauhari, R. (2019). Characterization of Bacterial Isolates found in Breeding Habitats of Mosquitoes in Dehradun City, Uttarakhand. International Journal of Advanced Research in Microbiology and Immunology, 1: 6-11.
Moreira, L. A., Iturbe-Ormaetxe, I., Jeffery, J. A., Lu, G., Pyke, A. T., Hedges, L. M., Rocha, B. C., Hall-Mendelin, S., Day, A. and Riegler, M. (2009). A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell, 139: 1268-1278.
Rajagopal, R. (2009). Beneficial interactions between insects and gut bacteria. Indian Journal of Microbiology, 49: 114-119.
Roy, M., Chatterjee, S., Roy, P. and Dangar, T. (2010). Significance of the midgut bacterium Pseudomonas fluorescens on Culex vishnui (Diptera: Culicidae) larval development. International Journal of Tropical Insect Science, 30: 182-185.
Shelomi, M. (2019). Bacterial and eukaryote microbiomes of mosquito habitats in dengue-endemic southern Taiwan. Journal of Asia-Pacific Entomology, 22: 471-480.
Smibert, R. and Krieg, N. (1995). Phenotypic testing. Methods for General and Molecular Bacteriology. American Society for Microbiology, 607-654.
Spencer, M., Blaustein, L. and Cohen, J. E. (2002). Oviposition habitat selection by mosquitoes (Culiseta longiareolata) and consequences for population size. Ecology, 83: 669-679.
Tamura, K., Dudley, J., Nei, M. and Kumar, S. (2007). MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular biology and evolution, 24: 1596-1599.
Technic, C. O. B., Pelczar, M. J., Bard, R. and Burnett, G. W. (1957). Manual of microbiological methods, McGraw-Hill Book Company.
Travanty, N. V., Apperson, C. S. and Ponnusamy, L. (2019). A diverse microbial community supports larval development and survivorship of the Asian tiger mosquito (Diptera: Culicidae). Journal of medical entomology, 56: 632-640.
Trexler, J. D., Apperson, C. S., Zurek, L., Gemeno, C., Schal, C., Kaufman, M., Walker, E., Watson, D. W. and Wallace, L. (2003). Role of Bacteria in Mediating the Oviposition Responses of Aedes albopictus (Diptera: Culicidae). Journal of Medical Entomology, 40: 841-848.
Why, A. M. and Walton, W. E. (2020). Oviposition Behavior of Culex tarsalis (Diptera: Culicidae) Responding to Semiochemicals Associated with the Western Mosquitofish, Gambusia affinis (Cyprinodontiformes: Poecilliidae). Journal of medical entomology, 57: 343-352.

