Ujjwal Malik![]()
Netaji Mahavidyalaya, Arambagh, Hooghly, West-Bengal, 712601, India
Corresponding author email: ujjwalvb@gmail.com
Article Publishing History
Received: 11/01/2025
Accepted After Revision: 15/03/2025
Opioid research has mostly been conducted on higher species and has hardly ever been documented in lower animals like the spiders, which are an excellent type species for examining behavior. The effects of opioids on the locomotor activity and exploratory behavior of an unknown area of the funnel-web spider Pireneitega luctuosa were examined along with the spontaneous behavior of spiders in an open-field setting using Software Intelligent 3.0 Behavior Tracker. Real-time movement was captured by high-resolution infrared cameras, and the software used movement trajectories to track important behavioral patterns like exploration, thigmotaxis, resting, and escape attempts. This automated technique allowed for a thorough evaluation of spider behavior by providing accurate, objective data on distance travelled, speed fluctuations, and zone preference.
The findings revealed the following: (a) The spontaneous behavior of spiders in the open field exhibited a high to low trend, meaning that they tended to be active in the early stages and quiet in the late stages, in the groups that received no treatment, distilled water treatment, and four different morphine concentration treatment groups. (b) The spider’s locomotor activity declined within 90 minutes of being exposed to opioid solutions, and there was a negative correlation between the morphine concentration and locomotor distance. (c) The varying morphine concentrations had no effect on the more careful exploration of the spider center region, which is more likely to be active in the periphery. (d) Following various opioid concentration treatments, the exploration duration and distance to the central region of the spider reduced, and the groups receiving distilled water and high concentration morphine solutions differed significantly. According to these present findings, opium not only temporarily stopped the funnel-web spider from acting spontaneously, but it also somewhat stopped the spider from exploring uncharted territory. The findings from this study provide valuable insights into the effects of opioids on arachnid behaviour, with potential applications in multiple fields.
Funnel-web spider; locomotor activities; exploration behavior; Opioid receptor agonist; Morphine
Malik U. Exposure to morphine Influences the Activity and Behavior of the Funnel-web Spider Pireneitega luctuosa. Biosc.Biotech.Res.Comm. 2025;18(1).
Malik U. Exposure to morphine Influences the Activity and Behavior of the Funnel-web Spider Pireneitega luctuosa. Biosc.Biotech.Res.Comm. 2024;18(1). Available from: <a href=”https://surl.li/mbagnb“>https://surl.li/mbagnb</a>
INTRODUCTION
Spiders belong to arena of arachnida of phylum arthropods,have a small brain size, and their nervous system is relatively simple compared with that of higher animals. But spiders are sensitive to external stimuli and move quickly, and through the long evolutionary process many groups of spiders have formed specific and complex behavior patterns, such as a lot of spiders use their web to hunt (Foelix 2011), some of spiders have extreme behavior-sexual cannibalism (Gavín-Centol et al; 2017, Fisher et al; 2018), one species of jumping spider have nursing behavior like higher animals (Chen et al; 2018).
The funnel-web spider is named for its three-dimensional funnel-shaped web, which is usually connected with the outside world through multiple entrances and exits. The funnel-web spider quietly lies at the main entrance, quickly attacks and captures prey caught by the web, and escapes from one of the exits quickly when encountering a predator. Previous reports and our studies have been found that sexual catalepsy behavior exists in many funnel-web spider species, that is, the female coward all the legs during mating for a long time, and the duration of sexual catalepsy of the spider of P. luctuosa can be up to 20 hours on average (Singer et al; 2000, Xiao et al; 2015, Liu et al; 2018, Frank, et al., 2023, Liu et al., 2024 ).
Thus it shows that the funnel-web spider is a class of spider groups with quick action and complex behavior, which is a good type animal for studying the behavior of lower animals.Morphine, a central nervous system drug, was first separated from opium in 1806 by the German chemist F.W.A Serturner, which mainly acts on μ-opioid receptors. Opioid receptors mediate a series of body reactions such as analgesia, hypnosis and respiratory depression, so they are widely used as strong analgesic clinic in medicine (Mercadante 2010, Torres-López et al; 2013). However, with the widespread use of morphine, its toxicity appear gradually, among which the most common is the physiological dependence and tolerance , as well as the resulting behavioral changes (Yayeh et al; 2016, Corder et al; 2018).
The research on neuro drugs such as morphine has been focused on higher animals, for example, the avoidance behavior of harmful stimuli in rats was significantly reduced after morphine treatment (Pahng et al; 2017), the rats showed severe anxious behavior after chronic morphine withdrawal (Kim et al; 2018). The effects of morphine on the lower animals have been little studied, and only a few insect species have been involved. For example, some studies have shown no significant difference in growth rate and developmental status between which feed on morphine carrion and those which does not feed on morphine carrion of the blow fly of Calliphora stygia (George et al; 2009). Honeybees (Apis mellifera) showed decreased on associative memory and motor activity after acute injection of morphine, while it with chronic treatment showed decreased associative memory, olfactory memory and behavioral activity, but increased tolerance to stimulus (Chen et al; 2014, Fu et al; 2013).
There are few studies on the behavior change of spider groups under nervous system drug stress, focus on the effect of drugs on spider web weaving behavior, such as Witt (1965) studied the two garden spider Araneus diadematus and Argiope aurantia the change of the weaving behavior after nerve drug treatment, the results showed that high concentrations of scopolamine was reduced weaving frequency, after Benzedrine treatment the processing screw spacing and irregularity of radius increases, weaving efficiency was reduced.
However caffeine not only increase the screw spacing and spider radius of irregularity, also reduces the area of the web (Hesselberg et al; 2004). The various behaviors that occur spontaneously after animals enter a new environment can reflect the mental state of the test animals, open field test is a good method to observe locomotor activity and explore behaviors of test animals, and is common used to evaluate the effects of neurological drugs on animal behaviors (Hull et al; 2004, El Hassani et al; 2005, Tremmel et al; 2012).
The total amount of locomotor activities of animals in the open field reflects the activity of their behaviors. The fear of the new environment leads to the activity of animals mainly in the periphery area of the open field, but the nature of exploration of animals force them to enter the central area ever and again. Some central stimulants can significantly increase locomotor activity in animals (Szopa et al; 2016), while certain antipsychotics can reduce exploratory behavior in animals (De Santis et al; 2016). In this study, Pireneitega luctuosa (L. Koch, 1878), a common funnel-web spider in southern China, was used to study the changes of the spider’s locomotor activities and exploratory behaviors after being exposed to morphine solutions.
Another study demonstrated that morphine exacerbates inflammatory responses and associated behavioral and hippocampal deficits in rodents, suggesting complex interactions between opioids and neural function (Ayieng’a et al., 2024). Some scientists report that chronic morphine administration in rats leads to distinctive behavioral changes, including decreased licking frequency, reduced water intake, and altered sleep posture (Yin et al., 2024).
MATERIAL AND METHODS
Collecting and raising of the funnel-web spider P. luctuosa: Spiderlings of the funnel-web spider P. luctuosa were collected and were reared in an air-conditioned laboratory with temperature 25 ± 2 ℃ and photoperiod 14 L: 10 D. The spiders were individually maintained in a transparent polypropylene box (14 cm L × 10 cm W × 5.5 cm H) with a small wet cotton on the bottle to supply drinking water and moisture for the spider(Schwartz, Elisabeth F., et al 2012). All the spiders were fed with 2-3 house flies (Musca domestica) twice a week. From time to time, fruit flies (Drosophila melanogaster) and flour beetle larvae (Tenebrio molitor) were supplied, respectively, as supplementary food.
Spiders in P. luctuosa generally molt 5-6 times during their lifespans. Juveniles in 3-4 instar, which weighed an average of 127.421 ± 11.259 g, were chosen for the experiment. All the chosen juveniles were fed with house flies one day before the experiment was conducted to eliminate possible undesirable effects.
Exposure of the spiders to morphine solutions: The powder-like morphine was diluted by distilled water into 4 types of concentrations: 7.5, 15, 30, and 60 mg/mL. The juvenile spiders chosen for the experiment were randomly assigned into 1 untreated group (without any treatment) and 5 treated groups, which were treated with distilled water and 4 morphine solutions in 7.5, 15, 30 and 60 mg/mL, respectively. The sample size for each group is 8 juvenile P. luctuosa. Distilled water and morphine solutions were contained in a 30 mL sprayer with a fine mist nozzle. All the treated juveniles were individually introduced into a 50 mL centrifuge tube and then sprayed by the corresponding distilled water or morphine solution. The amount of the distilled water or morphine solutions used for each juvenile was 0.1 mL. Then the treated spider was rested 10 min for the following examination of locomotor activity.
Behavioral test: The open field of the spider behavior test was a special round container made of polymethyl methacrylate (190 mm in diameter and 75 mm in height), which was used to record and analyze the weaving, hunting, courtship, mating and other behaviors (Liu et al; 2018) of funnel-web spiders (including P. luctuosa). Before each behavior test, the open field container was cleaned with anhydrous ethanol, then air-dried. Without treatment, distilled water treatment and different morphine concentration treatment groups of the spider of P. luctuosa were introduced into the open field; SMART 3.0 behavior tracker software (Panlab, Harvard Apparatus, and USA) was used to record the spontaneous behavior of the spider. In the software, the open field was divided into the central area and a periphery area according to the ratio of 3:1 in diameter; the distance and time of locomotor behavior in the central area and the periphery area were recorded. The data collection time was 2 h, and 8 individuals from each group.
Statistical Analysis: The distance and time of locomotor behavior of P. luctuosa were expressed as Mean ± SE, Linear fitting and Polynomial fitting analysis were used for the distance activities, the concentration of morphine in relation to the distance, distance changes over time, the Independent sample t-test was used to the compare of the distance and time of locomotor behavior in central area and periphery area. All statistical analyses were conducted using SPSS for Windows (version 19.0; IBM SPSS Statistics, USA), and graph was completed by Origin Pro (version 2018c, Origin Lab, USA).
Results
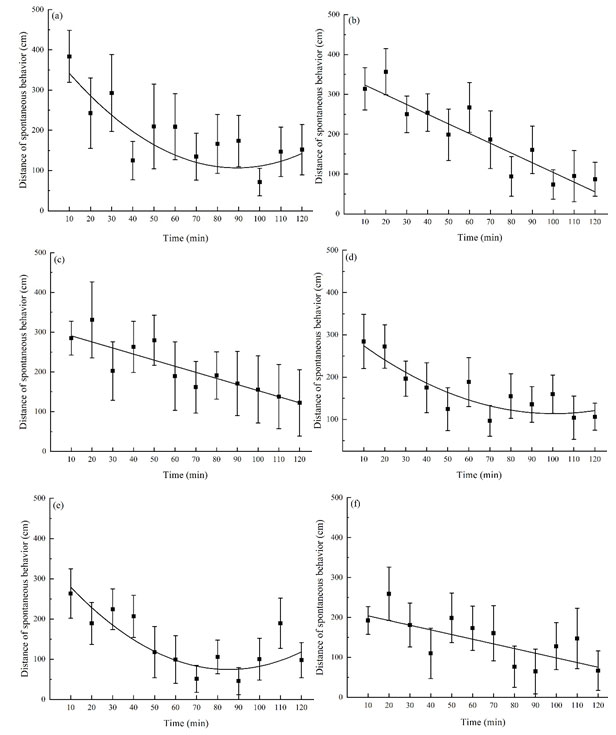
The locomotor activities of P. luctuosa in the open field changed with time after exposed to morphine solutions: Without treatment group of P. luctuosa was introduced into the open field for 10 minutes and became very active, crawling rapidly in succession along the periphery area of the open field. However, with the increase of time, continuous crawling turns into intermittent, and the crawling time gradually decreases, and the interval time becomes more. The last 30 minutes of the crawl were accompanied by the movement of touching the wall of the open field with the opening of the spinneret and the weaving, crossing the central area of the open field from time to time (Fig 1 a). After being treated with different concentrations of morphine (including the distilled water group), the locomotor distance of P. luctuosa within 10 minutes after being introduced into the open field was less than that of the without treatment group, but the overall trend was consistent, that is, the locomotor distance trend showed high to low (Fig 1 b-f).
Curve-fitting was performed for the locomotor activities of each group in open field, and the fitting equations were respectively: without treatment group y = 3.753x2 – 66.889x + 404.687, R2 = 0.601; distilled water group y = 347.694 – 24.368x, R2 = 0.862; 7.5 mg/mL group y = 306.096 – 15.318x, R2 = 0.779; 15 mg/mL group y = 1.956x2 – 39.360x + 311.811, R2 = 0.675; 30 mg/mL group y = 3.619x2 – 61.684x + 337.279, R2 = 0.650; 60 mg/mL group y = 215.711 – 11.689x, R2 = 0.561. (Fig 1 a-f).
Figure 1: Time-course of locomotor distance of P. luctuosa exposed to morphine solutions within 2 hours. The morphine concentrations were (a) 0 mg/mL (distilled water), (b) 7.5 mg/ml, (c) 15 mg/ml, (d) 30 mg/ml and (e) 60 mg/ml, respectively.
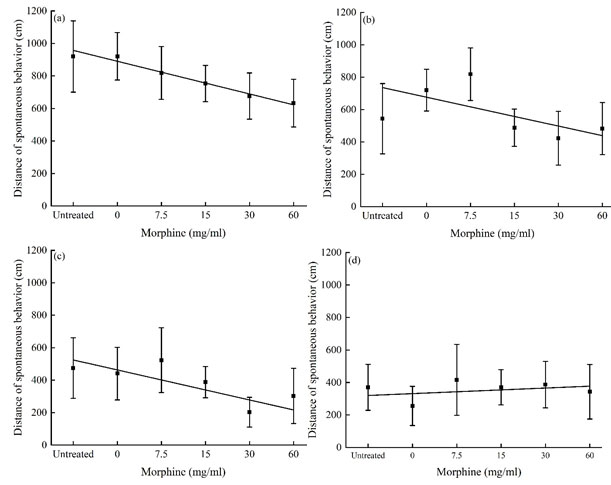
The difference in locomotor activities of P. luctuosa in the open field between groups with different time periods after exposed to morphine solutions: The total locomotor distance of each group was 2000-2500 cm during the 2 h monitoring period, the correlation between the locomotor distance and the concentration of morphine was analyzed, the results showed after exposed to morphine solutions, total locomotor distance of P. luctuosa within the first 30 min, 30-60 min and 60-90 min was negatively correlated with morphine concentration (Fig 2 a-c); However, during the last 30 min monitoring period, the locomotor distance of each group of P. luctuosa was basically the same, showing no relate of morphine concentration (Fig 2 d). The fitting equations were respectively: 30 min y = 1024.225 – 66.962x, R2 = 0.970; 30-60 min y = 795.241 – 59.467x, R2 = 0.414; 60-90 min y = 586.093 – 61.609x, R2 = 0.616; last 30 min y = 308.357 + 11.444x, R2 = 0.130. The data analysis results showed that within 90 min after exposed to morphine solutions, the locomotor distance of the spider was negatively correlated with morphine concentration. Indicating that morphine had a negative effect on the behavioral activity of the spider.
Figure 2: Time-course of locomotor distance of P. luctuosa exposed to morphine solutions in the 4 successive monitoring periods. (a) 0-30 min, (b) 30-60 min, (c) 60-90 min and (d) 90-120 min.
The area characteristics of locomotor activity in open field after exposed to morphine solutions: When the spider first was put into open field, showed crawling rapid succession along periphery area of the open field, with the increase of duration of stay in the open field, the spider of P. luctuosa began to explore the central area, which showed that every now and then, it quickly passed through the central area from the periphery area, or went to the central area and then returned to the periphery area, but the locomotor distance and the time spent in the central area are far from that in the peripheral area.
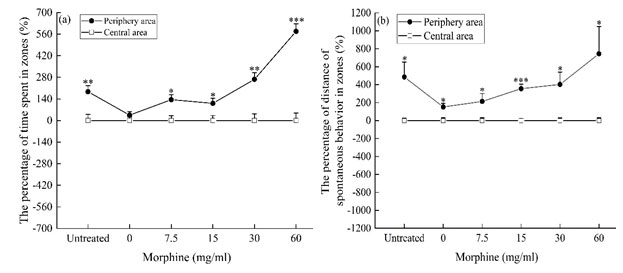
We compared and analyzed the locomotor activities of P. luctuosa in the central area and the periphery area, the duration of stay and locomotor distance of the central area were taken as the baseline, and the percentage chart of the duration of stay and locomotor distance of the peripheral area in the central area was plotting, that is (Time or distance of periphery area – mean time or distance of central area) / mean time or distance of central area × 100%. The results showed that during the 2-h monitoring period, except for the 0 mg/mL (distilled water) group (P > 0.05, Fig. 3a), the time spent in the central area of the untreated group, the 7.5 mg/mL group, the 15 mg/mL group, the 30 mg/mL group and the 60 mg/mL group were all significantly less than the time spent in the peripheral area (P < 0.05, Fig 3 a); The locomotor distance of all groups in the central area was less than that in the peripheral area (P < 0.05, Fig. 3b). The results indicated that the spider was cautious to explore the central area and tended to be active in the peripheral area, but the morphine solutions did not change the behavior tendency of the spider.
Figure 3: The percentage of distance and time of locomotor behavior in zones.
* P < 0.05, ** 0.001 < P < 0.01, *** P < 0.001. The duration of stay and locomotor distance of the central area were taken as the baseline, and the percentage chart of the duration of stay and locomotor distance of the peripheral area in the central area was plotting, that is (Time or distance of periphery area – mean time or distance of central area) / mean time or distance of central area ×100%.
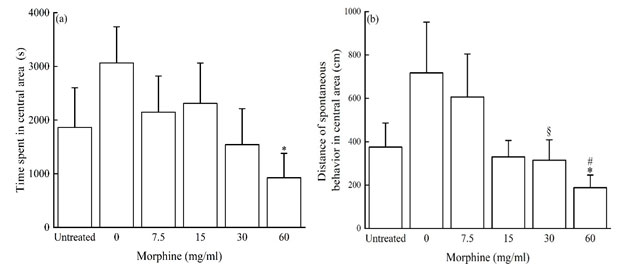
The difference of locomotor activity of P. luctuosa in central area of open field after exposed to morphine solutions between groups: We analyzed the inter-group differences in the exploration time and distance of P. luctuosa in the central area, the results showed that the 0 mg/mL (distilled water) group has the longest detention time in the central area (Fig. 4a) and the largest locomotor distance (Fig. 4b); After exposed to morphine solutions, the retention time and locomotor distance of P. luctuosa in the central area showed a decreasing trend, and the retention time in the central area of the 0 mg/mL (distilled water) group was significantly longer than that in the 60 mg/mL group, but the difference among other groups shows no significant (Fig. 4a). The locomotor distance in the central area of the 0 mg/mL (distilled water) group was significantly higher than that of the 30 mg/mL and 60 mg/mL groups, and that of the 7.5 g/mL group was also significantly higher than that of the 60 mg/mL group, with no significant difference among other groups (Fig. 4b). The results indicated that morphine inhibited spider’s exploration behavior to unknown areas to a certain extent, lead to its exploration time and distance to the unknown area decreased.
Figure 4: Distance of locomotion and time spent in central area in P. luctuosa exposed to morphine solutions within 2-hours. * means P < 0.05 vs. 0 mg/ml, § means P < 0.05 v.s.0 mg/ml, # means P < 0.05 vs. 7.5 mg/ml.
DISCUSSION
After being introduced into an open field, the locomotor activity in the early stage of monitoring was high, while in the later stage was low, and the weaving behaviors were rare and appeared in the late stage of monitoring, which may be related to the nature of spiders generally weaving webs at night (Ramousse et al; 1976), since our behavior experiments were conducted during the day, we rarely observed the weaving behavior of P. luctuosa that using end of abdomen (spinneret) to tap the inner wall of the open field; Like many of the higher animals tested (Lezak et al; 2017), P. luctuosa was mainly active in the peripheral area after entered the open field, and was cautious in exploring the central area, which was showed by rapid crossing of the central area or rapid retracing back to the peripheral area after reaching the central area. The behavior characteristics of P. luctuosa in the open field and its tendency to move in the peripheral area were not changed.
But, within 90min after exposed to morphine solutions, the locomotor distance of the spider decreased, and the distance was negatively correlated with morphine concentration; after exposed to morphine solutions, the exploration time and distance to the central area of P. luctuosa decreased, and the difference between the morphine treatment group and the 0 mg/mL (distilled water) group was significant. In this study, the spray treatment of the spider with a morphine solution or distilled water was similar to rainfall in the field. After being sprayed with distilled water, the spider’s exploration activity time and distance in the central area were the highest, which may be a stress response to the external rainfall.
In other words, after being stimulated by rainfall, the spider was more willing to explore the unknown area to seek shelter from the rain. However, the behavior of P. luctuosa after morphine solution spray became more cautious, especially after high concentration; its exploration time and distance were significantly lower than those in distilled water spray and low concentration of morphine solution groups. Hence, our results showed that morphine not only inhibited the behavioral activity of the spider in a certain period, but also inhibited the behavior of the spider in exploring unknown areas to a certain extent.
Morphine is an opioid receptor agonist with a variety of neuropsychopharmacological activities, which can act on the nervous system and affects animal behavior (Altarifi et al; 2015, Samuels et al; 2017, Roeckel et al; 2018). The spontaneous behavior of animals is a direct manifestation of the functional state of the central nervous system, and the activity of the corresponding nervous system increases when excited, and vice versa, so the locomotor activity of animals is an important indicator to evaluate the excited state of the central nervous system (Marques et al; 2014). Morphine treatment reduced locomotor activity of P. luctuosa in the new environment and inhibited its exploratory behavior, making it timid (preferring to stay on the peripheral area).
The reason may be that morphine has an inhibitory effect on the relevant functional areas of these invertebrate brain area, but the relevant physiological mechanism needs to be further studied by combining the methods of physiology, neurobiology, anatomy and other disciplines. We expect to explore the similarities and differences in the mechanism of behavior occurrence and neural regulation between higher animals and lower animals by studying the neurophysiological mechanism of complex behaviors in invertebrates such as spiders more extensively and more deeply.
Over the last few years, research has gone further and how morphine affects behavior and neural function in different animals, which can be useful for studying the funnel-web spider Pireneitega luctuosa. In 2024, a study established that morphine worsens inflammation and the resulting deterioration of hippocampal-dependent behavior and cognition in rodents, suggesting that opioids have a complicated relationship with the nervous system (Ayieng’a et al. 2024).
In 2024, a behavioral study how ever, found that rats given chronic morphine had several differences in their behavior, including less licking, drinking less water, and taking up a different sleeping position (Yin et al., 2024). There is hardly any research on spiders, particularly, P. luctuosa, directly, but studies of other invertebrates are helpful. For example, crayfish treated with morphine displayed increased activity and investigatory behavior, which was followed by withdrawal signs when the treatment was stopped, suggesting the existence of pathways that are sensitive to opioids and regulate activity.
Also, experiments on molluscs have established that morphine lengthens the reflex to noxious stimuli, and these effects are counteracted by opioid antagonists, thus suggesting that opioid receptors are evolved from very early organisms. These findings suggest that it is crucial to examine the impact of morphine on the activity and behaviour of P. luctuosa as it is likely to shed light on the basic mechanisms of opioid action in arachnids and may also help to further understand neurochemical control of behaviour in other organisms.
CONCLUSION
The varying morphine concentrations had no effect on the more careful exploration of the spider center region, which is more likely to be active in the periphery. Following various opioid concentration treatments, the exploration duration and distance to the central region of the spider were reduced, and the groups receiving distilled water and high concentration morphine solutions differed significantly. According to these present findings, opium not only temporarily stopped the funnel-web spider from acting spontaneously, but it also somewhat stopped the spider from exploring uncharted territory. The findings from this study provide valuable insights into the effects of opioids on arachnid behaviour, with potential applications in multiple fields.
Data Availability: All data will be made available on a reasonable request to the corresponding author.
Funding: Nil
Conflict of Interest: The author declares no conflict of interest.
REFERENCES
Altarifi A A, Rice K C, Negus S S. 2015. Effects of μ-Opioid Receptor Agonists in Assays of Acute Pain-Stimulated and Pain-Depressed Behavior in Male Rats: Role of μ-Agonist Efficacy and Noxious Stimulus Intensity. Journal of Pharmacology and Experimental Therapeutics, 352 (2): 208-217.
Ayieng’a, E.O., Afify, E.A., Abuiessa, S.A. et al. Morphine aggravates inflammatory, behavioral, and hippocampal structural deficits in septic rats. Sci Rep 13, 21460 (2023). https://doi.org/10.1038/s41598-023-46427-y
Christiansen A, Baum R, Witt P N. 1962. Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. The Journal of pharmacology and experimental therapeutics, 136(1): 31-37.
Chen Y M, Yu F, Yang H B, Yao T, Ma Y Y, Wang J H. 2014. Effects of Chronic Morphine Treatment on an Odor Conditioning Paradigm, Locomotor Activity and Sucrose Responsiveness in Honeybees (Apis mellifera). Journal of Insect Behavior, 27(6): 791-803.
Chen Z Q, Corlett R T, Jiao X G, Liu S J et al; 2018. Prolonged milk provisioning in a jumping spider. Science, 362 (6418): 1052-1055.
De Santis M, Lian J, Huang X F, Deng C. 2016. Early antipsychotic treatment in childhood/adolescent period has long-term effects on depressive-like, anxiety-like and locomotor behaviors in adult rats. Journal of Psychopharmacology, 30 (2): 204-214.
El Hassani A K, Dacher M, Gauthier M, Armengaud C. 2005. Effects of sub lethal doses of fipronil on the behavior of the honeybee (Apis mellifera). Pharmacology Biochemistry and Behavior, 82(1): 30-9.
Fu Y, Chen Y M, Yao T, Li P, Ma Y Y, Wang J H. 2013. Effects of morphine on associative memory and locomotor activity in honeybees (Apis mellifera). Neuroscience Bulletin, 29(3): 270-278.
Fisher A M, Cornell S J, Holwell G I, Price T A R. 2018. Sexual cannibalism and population viability. Ecology Evolution, 8(13): 6663-6670.
Frank SC , K. Christensen , R. Lourenc , D. Harms & B. A. Buzatto Mating behavior of the Sydney funnel‐web spider (Atracidae: Atrax robustus) and implications for the evolution of courtship in mygalomorph spiders.” Journal of Zoology 320.3 (2023): 169-178.
George K A, Archer M S, Green L M, Conlan X A, Toop T. 2009. Effect of morphine on the growth rate of Calliphora stygia (Fabricius) (Diptera: Calliphoridae) and possible implications for forensic entomology. Forensic Science International, 193(1): 21-25.
Gavín-Centol M P, Kralj-Fišer S, De Mas E,Ruiz-Lupión D, Moya-Laraño J. 2017. Feeding regime, adult age and sexual size dimorphism as determinants of pre-copulatory sexual cannibalism in virgin wolf spiders. Behavioral Ecology and Sociobiology, 71(1): 1-10.
Hull E M, Muschamp J W, Sato S. 2004. Dopamine and serotonin: influences on male sexual behavior. Physiology & behavior, 83(2): 291-307.
Hesselberg T, Vollrath F. 2004. The effects of neurotoxins on web-geometry and web-building behavior in Araneus diadematus Cl. Physiology & Behavior, 82(2): 519-529.
Kim D H, Kim N J, Zhao R J, Kim, D H, Yang C H, Kim H Y, et al; 2018. Effects of acupuncture on the anxiety-like behavior induced by withdrawal from chronic morphine use. Neuroscience letters, 664: 38-42.
Foelix R F. Biology of spiders. Third edition. Oxford: Oxford University Press, 2011: 155-187.
Lezak K R, Missig G, Carlezon Jr W A. 2017. Behavioral methods to study anxiety in rodents. Dialogues in Clinical Neuroscience, 19 (2): 181-191.
Liu J H, Xiao Y H, Chen Z W, Liu K K, Xu X. 2018. Comparative study on sexual behavior of three funnel-web spiders presenting catalepsy during copulation. Acta Arachnologica Sinica, 27 (1): 44-50.
Liu, Jihe, et al. “Females adopt sexual catalepsy to facilitate mating.” Current Zoology 70.2 (2024): 174-181.
Mercadante, S. 2010. Intravenous morphine for management of cancer pain. Lancet Oncology, 11(5): 484-489.
Marques H G, Bharadwaj A, Iida F. 2014. From spontaneous motor activity to coordinated behaviour: a developmental model. PLoS Computational Biology, 10(7): 1-20.
Pahng A R, Paulsen R I, Mcginn M A, Edwards, K N, & Edwards S. 2017. Neurobiological correlates of pain avoidance-like behavior in morphine-dependent and non-dependent rats. Neuroscience, 366: 1-14.
Rhodes M E, Creel T J, Nord A N. 2002. Sex differences in CNS neurotransmitter influences on behavior, in: Pfaff, D.W., Arnold, A.P., Fahrbach, S.E., Etgen, A.M., Rubin, R.T. (Eds.), Hormones, Brain and Behavior, Elsevier/Academic Press, San Diego, pp. 2747-2787.
Ramousse R, Davis III F. 1976. Web-building time in a spider: Preliminary applications of ultrasonic detection. Physiology & Behavior, 17 (6): 997-1000.
Roeckel L A, Massotte D, Olmstead M C, Befort K. 2018. CB1 Agonism Alters Addiction-Related Behaviors in Mice Lacking Mu or Delta Opioid Receptors. Frontiers in Psychiatry, 9: 1-10.
Samuels B, Nautiyal K, Kruegel A. et al; 2017. The Behavioral Effects of the Antidepressant Tianeptine Require the Mu-Opioid Receptor. Neuropsychopharmacol 42: 2052-2063.
Seidel E R, Beaton J M, Teague R S. 1979. The effects of cholinergic agents on morphine-induced circling behavior in the intact mouse. European Journal of Pharmacology, 56(1–2): 75-80.
Singer F, Riechert S E, Xu H, Morris A W, Becker E, Hale J A , Noureddine M A. 2000. Analysis of courtship success in the funnel-web spider Agelenopsis aperta. Behaviour, 137(1): 93-117.
Schwartz EF, Caroline B F Mourão, Karla G Moreira, Thalita S Camargos, Márcia R Mortari (2012) Arthropod venoms: a vast arsenal of insecticidal neuropeptides.” Peptide science 98.4 : 385-405.
Szopa A, Poleszak E, Wyska E. et al; 2016. Caffeine enhances the antidepressant-like activity of common antidepressant drugs in the forced swim test in mice. Naunyn-Schmiedeberg’s Archives Pharmacology, 389 (2): 211-221.
Tremmel M, Müller C. 2012. Insect personality depends on environmental conditions. Behavioral Ecology, 24(2): 386-92.
Torres-López J E, Carmona-Díaz E, Cortés-Peñaloza J L, Guzmán-Priego C G, & Rocha-González H I. 2013. Antinociceptive synergy between diclofenac and morphine after local injection into the inflamed site. Pharmacological reports: PR, 65(2): 358-367.
Witt P N, Reed C F. 1965. Spider-web building. Science, 149(3689): 1190.
Xiao Y H, Zunic-Kosi A, Zhang L W, Prentice T R, et al; 2015. Male adaptations to minimize sexual cannibalism during reproduction in the funnel-web spider Hololena curta. Insect science, 22(6): 840-852.
Yayeh T, Yun K, Jang S, Oh S. 2016. Morphine dependence is attenuated by red ginseng extract and ginsenosides rh2, rg3, and compound k. Journal of Ginseng Research, 40(4): 445-452.
Yin, Jie, et al. “Behavioral video coding analysis of chronic morphine administration in rats.” Biomedical Reports 21.5 (2024): 168. Yin, Jie, et al. “Behavioral video coding analysis of chronic morphine administration in rats.” Biomedical Reports 21.5 (2024): 168.
Zhan H, Tang G X, Xin Y M. 1999. Dose and temporal features of the effects of several central stimulants and hypnotics on locomotor activity in mice. Chin J Aerospace Med, 10(3): 133-137.
Zhao S S, Zhang K X, Bao S H, Ye Q Q, Wang D, Chen D S, et al; 2013. Effects of protocol on spontaneous motor activity and learning behaviors in juvenile mice. Chinese Pharmacological Bulletin, 29(2): 198-201.

